Κυκλινο-εξαρτώμενη κινάση
Οι κυκλινο-εξαρτώμενες κινάσες ή κινάσες εξαρτώμενες από κυκλίνη (Cyclin-dependent kinases, CDK) είναι οι οικογένειες πρωτεϊνικών κινασών που ανακαλύφθηκαν για πρώτη φορά λόγω του ρόλου τους στη ρύθμιση του κυτταρικού κύκλου. Συμμετέχουν επίσης στη ρύθμιση της μεταγραφής, στην επεξεργασία του mRNA και στη διαφοροποίηση των νευρικών κυττάρων.[1] Είναι παρούσες σε όλους τους γνωστούς ευκαρυώτες και η ρυθμιστική τους λειτουργία στον κυτταρικό κύκλο έχει διατηρηθεί εξελικτικά. Τα κύτταρα ζύμης μπορούν μάλιστα να πολλαπλασιαστούν κανονικά όταν το CDK γονίδιό τους έχει αντικατασταθεί με το ομόλογο ανθρώπινο γονίδιο. [2] Οι CDK είναι σχετικά μικρές πρωτεΐνες, με μοριακά βάρη που κυμαίνονται από 34 έως 40 kDa, και δεν περιέχουν πολλά περισσότερα εκτός της επικράτειας κινάσης.

Εξ ορισμού, μια CDK δεσμεύεται με μια ρυθμιστική πρωτεΐνη που ονομάζεται κυκλίνη. Χωρίς κυκλίνη, η CDK έχει μικρή δραστικότητα κινάσης. Μόνο το σύμπλοκο κυκλίνης-CDK είναι μια ενεργή κινάση. Οι CDK φωσφορυλιώνουν τα υποστρώματά τους σε σερίνες και θρεονίνες, επομένως συνιστούν κινάσες σερίνης-θρεονίνης. Η συναινετική αλληλουχία για τη θέση φωσφορυλίωσης στην αμινοξική αλληλουχία ενός υποστρώματος CDK είναι η [S/T*]PX[K/R], όπου S/T* είναι η φωσφορυλιωμένη σερίνη ή θρεονίνη, Ρ είναι προλίνη, Χ είναι οποιοδήποτε αμινοξύ, Κ είναι η λυσίνη και R είναι η αργινίνη.
Τύποι
Επεξεργασία| CDK | Συνοδή κυκλίνη | Λειτουργία | Ελλειμματικός φαινότυπος σε ποντίκια |
|---|---|---|---|
| Cdk1 | Κυκλίνη Β | Φάση Μ | Κανένας |
| Cdk2 | Κυκλίνη Ε | Μετάβαση G1 / S | Μειωμένο μέγεθος, προκαλεί πολλαπλασιασμό νευρικών προγονικών κυττάρων. Βιώσιμα, αλλά τόσο τα αρσενικά όσο και τα θηλυκά είναι στείρα |
| Cdk2 | Κυκλίνη Α | Φάση S, φάση G2 | |
| Cdk3 | Κυκλίνη C | Φάση G1 | Χωρίς ελαττώματα. Βιώσιμα, γόνιμα. |
| Cdk4 | Κυκλίνη D | Φάση G1 | Μειωμένο μέγεθος, διαβήτης λόγω έλλειψης ινσουλίνης. Βιώσιμα, αλλά τόσο τα αρσενικά όσο και τα θηλυκά είναι στείρα |
CDK και κυκλίνες στον κυτταρικό κύκλο
ΕπεξεργασίαΤα περισσότερα από τα γνωστά σύμπλοκα κυκλίνης-CDK ρυθμίζουν την πρόοδο του κυτταρικού κύκλου. Τα ζωικά κύτταρα περιέχουν τουλάχιστον εννέα CDK, τέσσερις εκ των οποίων, οι CDK1, 2, 3 και 4, εμπλέκονται άμεσα στη ρύθμιση του κυτταρικού κύκλου.[1] Σε κύτταρα θηλαστικών, η CDK1, με τις συνοδές κυκλίνες Α2 και Β1, μόνη της μπορεί να προωθήσει τον κυτταρικό κύκλο. [4] Μια άλλη, η CDK7, εμπλέκεται έμμεσα ως ενεργοποιητική κινάση της CDK. Τα σύμπλοκα κυκλίνης-CDK φωσφορυλιώνουν υποστρώματα κατάλληλα για την εκάστοτε φάση του κυτταρικού κύκλου. [3] Τα σύμπλοκα κυκλίνης-CDK προηγούμενων φάσεων του κυτταρικού κύκλου βοηθούν στην ενεργοποίηση συμπλόκων κυκλίνης-CDK σε μεταγενέστερη φάση.
| Φάση | Κυκλίνη | CDK |
|---|---|---|
| G0 | C | Cdk3 |
| G1 | D, E | Cdk4, Cdk2, Cdk6 |
| S | Α, Ε | Cdk2 |
| G2 | A | Cdk2, Cdk1 |
| Μ | B | Cdk1 |
Ρύθμιση δραστηριότητας
ΕπεξεργασίαΤα επίπεδα των κυκλινο-εξαρτώμενων κινασών (CDK) παραμένουν σχετικά σταθερά καθ 'όλη τη διάρκεια του κυτταρικού κύκλου και η ρύθμιση είναι κατά βάση μετα-μεταφραστική. Οι περισσότερες πληροφορίες για τη δομή και τη λειτουργία των CDK προέρχονται από τις CDK του S. pombe (Cdc2), του S. cerevisiae (CDC28) και των σπονδυλωτών (CDC2 και CDK2). Οι τέσσερις κύριοι μηχανισμοί ρύθμισης των CDK είναι η πρόσδεση κυκλίνης, η φωσφορυλίωση από CAK, η ρυθμιστική ανασταλτική φωσφορυλίωση και η σύνδεση αναστολέων των CDK (CKI). [5]
Πρόσδεση κυκλίνης
ΕπεξεργασίαΤο ενεργό κέντρο, ή η θέση δέσμευσης ΑΤΡ, όλων των κινασών είναι μια κοιλότητα μεταξύ ενός μικρού αμινο-τελικού λοβού και ενός μεγαλύτερου καρβοξυ-τελικού λοβού.[1] Η δομή της ανθρώπινης Cdk2 αποκάλυψε ότι οι CDK διαθέτουν μια τροποποιημένη θέση πρόσδεσης ΑΤΡ που μπορεί να ρυθμιστεί με πρόσδεση κυκλίνης. Η φωσφορυλίωση από μια ενεργοποιητική κινάση της CDK (CAK) στη θέση Thr 161 στον Τ-βρόχο αυξάνει την δραστηριότητα του συμπλόκου. Χωρίς κυκλίνη, ένας εύκαμπτος βρόχος που ονομάζεται βρόχος ενεργοποίησης ή βρόχος Τ μπλοκάρει την κοιλότητα και η θέση πολλών βασικών καταλοίπων αμινοξέων δεν είναι η βέλτιστη για πρόσδεση ΑΤΡ. Με την κυκλίνη, δύο α-έλικες αλλάζουν θέση για να επιτρέψουν τη σύνδεση ATP. Ένας από αυτούς, η έλικα L12 που βρίσκεται λίγο πριν από τον βρόχο T στην κύρια αλληλουχία, γίνεται ένας β-κλώνος και βοηθά στην αναδιάταξη του βρόχου T, ώστε να μην αποκλείει πλέον το ενεργό κέντρο. Η άλλη α-έλικα που ονομάζεται έλικα PSTAIRE αναδιατάσσεται και βοηθά στην αλλαγή της θέσης των βασικών καταλοίπων αμινοξέων στο ενεργό κέντρο.
Υπάρχει σημαντική εξειδίκευση αναφορικά με το ποια κυκλίνη συνδέεται με ποια CDK. [3] Επίσης, η πρόσδεση κυκλίνης καθορίζει την ειδικότητα του συμπλόκου κυκλίνης-CDK για συγκεκριμένα υποστρώματα. Οι κυκλίνες μπορούν να συνδέσουν απευθείας το υπόστρωμα ή να περιορίσουν τη CDK σε μια υποκυτταρική περιοχή όπου βρίσκεται το υπόστρωμα. Η εξειδίκευση υποστρώματος των S κυκλινών προσδίδεται από την υδρόφοβη ομάδα (με επίκεντρο την αλληλουχία MRAIL), η οποία έχει συνάφεια για πρωτεΐνες υποστρώματος που περιέχουν ένα υδρόφοβο RXL (ή Cy) μοτίβο. Οι κυκλίνες Β1 και Β2 μπορούν να περιορίσουν τη Cdk1 στον πυρήνα και στο Golgi αντίστοιχα, μέσω μιας ακολουθίας εντοπισμού εκτός της περιοχής δέσμευσης CDK. [1]
Φωσφορυλίωση
Επεξεργασία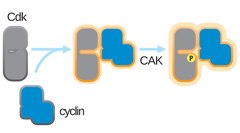
Η πλήρης δραστικότητα κινάσης της CDK απαιτεί ενεργοποιητική φωσφορυλίωση σε μια θρεονίνη δίπλα στο ενεργό της κέντρο.[1] Η ταυτότητα της ενεργοποιητικής κινάσης της CDK (CAK) που εκτελεί αυτήν τη φωσφορυλίωση ποικίλλει μεταξύ των οργανισμών-μοντέλων. Η χρονική στιγμή αυτής της φωσφορυλίωσης ποικίλλει επίσης. Σε κύτταρα θηλαστικών, η ενεργοποιητική φωσφορυλίωση συμβαίνει μετά τη πρόσδεση κυκλίνης. Στα κύτταρα ζύμης, εμφανίζεται πριν από τη σύνδεση της κυκλίνης. Η δραστικότητα των CAK δεν ρυθμίζεται από γνωστές οδούς κυτταρικού κύκλου και η πρόσδεση κυκλίνης είναι το περιοριστικό βήμα για την ενεργοποίηση της CDK.
Σε αντίθεση με την ενεργοποιητική φωσφορυλίωση, η ανασταλτική φωσφορυλίωση των CDK είναι ζωτικής σημασίας για τη ρύθμιση του κυτταρικού κύκλου. Διάφορες κινάσες και φωσφατάσες ρυθμίζουν την κατάσταση φωσφορυλίωσης τους. Μία από τις κινάσες που φωσφορυλιώνουν τυροσίνη είναι η Wee1, μια συντηρημένη σε όλους τους ευκαρυώτες κινάση.[1] Ο σχιζοσακχαρομύκητας (S. pombe) περιέχει επίσης μια δεύτερη κινάση Mik1 που μπορεί να φωσφορυλιώσει την τυροσίνη. Τα σπονδυλωτά περιέχουν μια διαφορετική δεύτερη κινάση που ονομάζεται Myt1 και σχετίζεται με τη Wee1 αλλά μπορεί να φωσφορυλιώσει τόσο τη θρεονίνη όσο και την τυροσίνη. Οι φωσφατάσες της οικογένειας Cdc25 αποφωσφορυλιώνουν τόσο τη θρεονίνη όσο και την τυροσίνη.
Αναστολείς των CDK
ΕπεξεργασίαΈνας αναστολέας κυκλινο-εξαρτώμενων κινασών (CKI) είναι μια πρωτεΐνη που αλληλεπιδρά με ένα σύμπλοκο κυκλίνης-CDK για την παρεμπόδιση της δραστικότητας κινάσης, συνήθως κατά τη διάρκεια της φάσης G1 ή σε απόκριση σε σήματα από το περιβάλλον ή από κατεστραμμένο DNA. [1] Στα ζωικά κύτταρα, υπάρχουν δύο μεγάλες οικογένειες CKI: η οικογένεια INK4 και η οικογένεια CIP/KIP. Οι πρωτεΐνες της οικογένειας INK4 είναι αυστηρά ανασταλτικές και δεσμεύουν μονομερή CDK. Οι κρυσταλλικές δομές των συμπλοκών CDK6-INK4 δείχνουν ότι η σύνδεση ΙΝΚ4 περιστρέφει τη CDK ώστε να μεταβάλλει τη δέσμευση κυκλίνης και τη δραστικότητα κινάσης. Οι πρωτεΐνες της οικογένειας CIP/KIP δεσμεύουν τόσο την κυκλίνη όσο και την CDK ενός συμπλόκου και μπορεί να είναι ανασταλτικές ή ενεργοποιητικές. Οι πρωτεΐνες της οικογένειας CIP/KIP ενεργοποιούν τα σύμπλοκα της κυκλίνης D με CDK4 ή CDK6 ενισχύοντας τον σχηματισμό συμπλόκου.
Στη ζύμη και τη Drosophila, οι CKI είναι ισχυροί αναστολείς των S- και M-CDK, αλλά δεν αναστέλλουν τις G1/S-CDK. Κατά τη διάρκεια της φάσης G1, υψηλά επίπεδα CKI εμποδίζουν την εμφάνιση γεγονότων του κυτταρικού κύκλου με λανθασμένη σειρά, αλλά δεν εμποδίζουν τη μετάβαση από το σημείο ελέγχου Έναρξη, το οποίο ξεκινά μέσω των G1/S-CDK. Μόλις ξεκινήσει ο κυτταρικός κύκλος, η φωσφορυλίωση από πρώιμες G1/S-CDK οδηγεί σε καταστροφή των CKI, αποτρέποντας την αναστολή σε μεταγενέστερες μεταβάσεις του κυτταρικού κύκλου. Στα κύτταρα θηλαστικών, η ρύθμιση των CKI λειτουργεί διαφορετικά. Η πρωτεΐνη p27 (Dacapo στη Drosophila) αναστέλλει τις G1/S- και S-CDK, αλλά δεν αναστέλλει τις S- και M-CDK. [1]
Suk1 ή Cks
ΕπεξεργασίαΟι CDK που εμπλέκονται άμεσα στη ρύθμιση του κυτταρικού κύκλου συνδέονται με μικρές πρωτεΐνες από 9 έως 13 kDa που ονομάζονται Suk1 ή Cks. [3] Αυτές οι πρωτεΐνες απαιτούνται για τη λειτουργία των CDK, αλλά ο ακριβής ρόλος τους είναι άγνωστος. Η Cks1 δεσμεύει τον καρβοξυ-λοβό της CDK και αναγνωρίζει φωσφορυλιωμένα κατάλοιπα. Μπορεί να βοηθά το σύμπλοκο κυκλίνης-CDK με υποστρώματα που έχουν πολλαπλές θέσεις φωσφορυλίωσης αυξάνοντας τη συγγένεια για το υπόστρωμα.
Άλλοι ενεργοποιητές
ΕπεξεργασίαΙικές κυκλίνες
ΕπεξεργασίαΟι ιοί μπορούν να κωδικοποιήσουν πρωτεΐνες με αλληλουχία ομόλογη των κυκλινών. Ένα καλά μελετημένο παράδειγμα είναι η Κ-κυκλίνη (ή v-κυκλίνη) από τον ερπητοϊό του σαρκώματος Kaposi (βλ. σάρκωμα Kaposi), που ενεργοποιεί τη CDK6. Τα σύμπλοκα ιικής κυκλίνης-CDK έχουν διαφορετικές ειδικότητες υποστρώματος και ευαισθησίες ρύθμισης. [6]
Ενεργοποιητές της CDK5
ΕπεξεργασίαΟι πρωτεΐνες p35 και p39 ενεργοποιούν τη CDK5. Αν και δεν έχουν ομόλογη αλληλουχία κυκλίνης, οι κρυσταλλικές δομές δείχνουν ότι η p35 αναδιπλώνεται με παρόμοιο τρόπο όπως οι κυκλίνες. Ωστόσο, η ενεργοποίηση της CDK5 δεν απαιτεί φωσφορυλίωση βρόχου ενεργοποίησης. [6]
RINGO/Speedy
ΕπεξεργασίαΠρωτεΐνες χωρίς ομολογία με την οικογένεια των κυκλινών μπορούν να είναι άμεσοι ενεργοποιητές των CDK. [7] Μια οικογένεια τέτοιων ενεργοποιητών είναι η οικογένεια RINGO/Speedy, η οποία αρχικά ανακαλύφθηκε στον Xenopus. Και τα πέντε μέλη που ανακαλύφθηκαν μέχρι σήμερα ενεργοποιούν άμεσα τις Cdk1 και Cdk2, αλλά το σύμπλοκο RINGO/Speedy-CDK2 αναγνωρίζει διαφορετικά υποστρώματα από το σύμπλοκο κυκλίνης A-CDK2.[6]
Παραπομπές
Επεξεργασία- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 1,6 1,7 Morgan, David O. (2007). The Cell Cycle: Principles of Control (1st έκδοση). London: New Science Press. ISBN 978-0-87893-508-6.
- ↑ «Complementation used to clone a human homologue of the fission yeast cell cycle control gene cdc2». Nature 327 (6117): 31–5. 1987. doi:. PMID 3553962.
- ↑ 3,0 3,1 3,2 3,3 «Cyclin-dependent kinases: engines, clocks, and microprocessors». Annual Review of Cell and Developmental Biology 13: 261–91. 1997. doi:. PMID 9442875. https://archive.org/details/sim_annual-review-of-cell-and-developmental-biology_1997_13/page/261.
- ↑ 4,0 4,1 «Mammalian cell-cycle regulation: several Cdks, numerous cyclins and diverse compensatory mechanisms». Oncogene 28 (33): 2925–39. August 2009. doi:. PMID 19561645.
- ↑ «Principles of CDK regulation». Nature 374 (6518): 131–4. March 1995. doi:. PMID 7877684.
- ↑ 6,0 6,1 6,2 «CDK activation by non-cyclin proteins». Current Opinion in Cell Biology 18 (2): 192–8. April 2006. doi:. PMID 16488127.
- ↑ «RINGO C is required to sustain the spindle-assembly checkpoint». Journal of Cell Science 123 (Pt 15): 2586–95. August 2010. doi:. PMID 20605920.