Ωχρό σωμάτιο
Το ωχρό σωμάτιο (λατινικά: corpus luteum, σημαίνει «κίτρινο σώμα») είναι μια προσωρινή ενδοκρινής δομή στις γυναικείες ωοθήκες και εμπλέκεται στην παραγωγή σχετικά υψηλών επιπέδων προγεστερόνης και μέτριων επιπέδων οιστραδιόλης και αναστολίνης Α.[1][2] Είναι τα υπολείμματα του ωοθυλακίου των ωοθηκών που απελευθέρωσαν ένα ώριμο ωάριο κατά τη διάρκεια μιας προηγούμενης ωορρηξίας.[3]

Το ωχρό σωμάτιο χρωματίζεται ως αποτέλεσμα της συγκέντρωσης καροτενοειδών (συμπεριλαμβανομένης της λουτεΐνης) από τη διατροφή και εκκρίνει μια μέτρια ποσότητα οιστρογόνων που αναστέλλει την περαιτέρω απελευθέρωση της ορμόνης απελευθέρωσης γοναδοτροπίνης (GnRH) και επομένως την έκκριση της ωχρινοτρόπου ορμόνης (LH) και της ωοθυλακιοτρόπου ορμόνη (FSH). Ένα νέο ωχρό σωμάτιο αναπτύσσεται με κάθε εμμηνορροϊκό κύκλο.
Ανάπτυξη και δομή
ΕπεξεργασίαΤο ωχρό σωμάτιο αναπτύσσεται από ένα ωοθυλάκιο κατά τη διάρκεια της ωχρινικής φάσης του εμμηνορροϊκού κύκλου ή του οιστρικού κύκλου, μετά την απελευθέρωση ενός δευτερεύοντος ωαρίου από το ωοθυλάκιο κατά την ωορρηξία. Το ωοθυλάκιο σχηματίζει πρώτα ένα ερυθρό (αιμορραγικό) σώμα προτού γίνει ωχρό σωμάτιο, αλλά ο όρος αναφέρεται στην ορατή συλλογή αίματος, που φαίνεται μετά τη ρήξη του ωοθυλακίου, που εκκρίνει προγεστερόνη. Ενώ το ωοκύτταρο (αργότερα ο ζυγώτης αν συμβεί γονιμοποίηση) διασχίζει τη σάλπιγγα στη μήτρα, το ωχρό σωμάτιο παραμένει στην ωοθήκη.
Το ωχρό σωμάτιο είναι συνήθως πολύ μεγάλο σε σχέση με το μέγεθος της ωοθήκης. Στους ανθρώπους, το μέγεθος της δομής κυμαίνεται από λιγότερο από 2 cm έως 5 cm σε διάμετρο.[4]
Τα κύτταρά του αναπτύσσονται από τα ωοθυλακικά κύτταρα που περιβάλλουν το ωοθυλάκιο της ωοθήκης.[5] Τα θυλακιώδη κύτταρα της θήκης ωχρινοειδοποιούνται σε μικρά ωχρινικά κύτταρα και τα θυλακιώδη κοκκιώδη κύτταρα σε μεγάλα ωχρινικά κύτταρα σχηματίζοντας το ωχρό σωμάτιο. Η προγεστερόνη συντίθεται από τη χοληστερόλη τόσο από τα μεγάλα όσο και από τα μικρά ωχρινικά κύτταρα κατά την ωχρινική ωρίμανση. Τα σύμπλοκα χοληστερόλης- LDL συνδέονται με υποδοχείς στην πλασματική μεμβράνη των ωχρινικών κυττάρων και μεταφέροντάς τα στο εσωτερικό του κυττάρου. Η χοληστερόλη απελευθερώνεται και αποθηκεύεται μέσα στο κύτταρο ως εστέρας χοληστερόλης. Η LDL ανακυκλώνεται για περαιτέρω μεταφορά χοληστερόλης. Τα μεγάλα ωχρινικά κύτταρα παράγουν περισσότερη προγεστερόνη λόγω των μη ανασταλμένων/βασικών επιπέδων δραστηριότητας της πρωτεϊνικής κινάσης Α (PKA) εντός του κυττάρου. Τα μικρά ωχρινικά κύτταρα έχουν υποδοχείς LH που ρυθμίζουν τη δραστηριότητα PKA εντός του κυττάρου. Η PKA φωσφορυλιώνει ενεργά τη στεροειδογόνο οξεία ρυθμιστική πρωτεΐνη (StAR) και την πρωτεΐνη μετατοπιστή για τη μεταφορά της χοληστερόλης από την εξωτερική μιτοχονδριακή μεμβράνη στην εσωτερική μιτοχονδριακή μεμβράνη.[6]
Η ανάπτυξη του ωχρού σωματίου συνοδεύεται από αύξηση του επιπέδου του στεροειδογόνου ενζύμου P450scc που μετατρέπει τη χοληστερόλη σε πρεγνενολόνη στα μιτοχόνδρια.[7] Η πρεγνενολόνη στη συνέχεια μετατρέπεται σε προγεστερόνη που εκκρίνεται από το κύτταρο και στην κυκλοφορία του αίματος. Κατά τη διάρκεια του οιστρικού κύκλου των βοοειδών, τα επίπεδα της προγεστερόνης στο πλάσμα αυξάνονται παράλληλα με τα επίπεδα του P450scc και της αδρενοδοξίνης του δότη ηλεκτρονίων, υποδεικνύοντας ότι η έκκριση προγεστερόνης είναι αποτέλεσμα ενισχυμένης έκφρασης του P450scc στο ωχρό σωμάτιο.[7]
Η αλυσίδα μεταφοράς ηλεκτρονίων του μιτοχονδριακού συστήματος P450, συμπεριλαμβανομένης της αναγωγάσης της αδρενοδοξίνης και της αδρενοδοξίνης, έχει αποδειχθεί ότι διαρρέει ηλεκτρόνια που οδηγούν στο σχηματισμό ρίζας υπεροξειδίου.[8][9] Προφανώς για να αντιμετωπιστούν οι ελεύθερες ρίζες που παράγονται από αυτό το σύστημα και από τον ενισχυμένο μιτοχονδριακό μεταβολισμό, τα επίπεδα των αντιοξειδωτικών ενζύμων καταλάση και υπεροξειδικής δισμουτάσης αυξάνονται επίσης παράλληλα με την ενισχυμένη στεροειδογένεση στο ωχρό σωμάτιο.[7]
| Θυλακική δομή | Ωχρινική δομή | Εκκριση |
| Κύτταρα θήκης | Ωχρινικά κύτταρα θήκης | ανδρογόνα,[10] προγεστερόνη[10] |
| Κοκκιώδη κύτταρα | Κοκκιώδη ωχρινικά κύτταρα | προγεστερόνη,[5] οιστρογόνα (πλειοψηφία),[5] και αναστολίνη Α[5][10] |
Όπως και τα προηγούμενα κύτταρα θήκης, τα ωχρινικά κύτταρα θήκης δεν διαθέτουν το ένζυμο αρωματάση που είναι απαραίτητο για την παραγωγή οιστρογόνων, επομένως μπορούν να κάνουν στεροειδογένεση μόνο μέχρι το σχηματισμό ανδρογόνων. Τα κοκκιώδη ωχρινικά κύτταρα έχουν όντως αρωματάση και τη χρησιμοποιούν για να παράγουν οιστρογόνα, χρησιμοποιώντας τα ανδρογόνα που είχαν συντεθεί προηγουμένως από τα ωχρινικά κοκκιώδη κύτταρα, καθώς τα κοκκιώδη ωχρινικά κύτταρα από μόνα τους δεν έχουν την 17α-υδροξυλάση ή την 17,20 λυάση για να παράγουν ανδρογόνα.[5] Μόλις το ωχρό σωμάτιο υποστραφεί, το υπόλειμμα είναι γνωστό ως λευκό σωμάτιο.[11]
Λειτουργία
ΕπεξεργασίαΤο ωχρό σωμάτιο είναι απαραίτητο για τη δημιουργία και τη διατήρηση της εγκυμοσύνης στις γυναίκες. Το ωχρό σωμάτιο εκκρίνει προγεστερόνη, η οποία είναι μια στεροειδής ορμόνη υπεύθυνη για την ανάπτυξη του ενδομητρίου και τη συντήρησή του, αντίστοιχα. Παράγει επίσης ρελαξίνη, μια ορμόνη που είναι υπεύθυνη για το μαλάκωμα της ηβικής σύμφυσης που βοηθά στον τοκετό.
Ανεπιτυχής γονιμοποίηση
ΕπεξεργασίαΕάν το ωάριο δεν γονιμοποιηθεί, το ωχρό σωμάτιο σταματά να εκκρίνει προγεστερόνη και υποστρέφεται (μετά από περίπου 10 ημέρες στον άνθρωπο). Στη συνέχεια εκφυλίζεται σε λευκό σωμάτιο, το οποίο είναι μια μάζα ινώδους ουλώδους ιστού.
Η εσωτερική επένδυση της μήτρας (ενδομήτριο) αποβάλλεται μέσω του κόλπου (σε θηλαστικά που περνούν από έναν εμμηνορροϊκό κύκλο ). Σε έναν οιστρικό κύκλο, το ενδομήτριο εκφυλίζεται ξανά στο κανονικό μέγεθος.
Επιτυχής γονιμοποίηση
Επεξεργασία
Εάν το ωάριο γονιμοποιηθεί και συμβεί εμφύτευση, τα κύτταρα συγκυτιοτροφοβλάστης (που προέρχονται από την τροφοβλάστη) της βλαστοκύστης εκκρίνουν την ορμόνη ανθρώπινη χοριακή γοναδοτροπίνη (hCG, ή παρόμοια ορμόνη σε άλλα είδη) την 9η ημέρα μετά τη γονιμοποίηση.
Η ανθρώπινη χοριακή γοναδοτροπίνη σηματοδοτεί το ωχρό σωμάτιο να συνεχίσει την έκκριση προγεστερόνης, διατηρώντας έτσι την παχιά επένδυση (ενδομήτριο) της μήτρας και παρέχοντας μια περιοχή πλούσια σε αιμοφόρα αγγεία στην οποία μπορεί να αναπτυχθεί ο ζυγώτης.
Η εισαγωγή προσταγλανδινών σε αυτό το σημείο προκαλεί εκφύλιση του ωχρού σωματίου και αποβολή του εμβρύου. Ωστόσο, σε ζώα με πλακούντα όπως ο άνθρωπος, ο πλακούντας τελικά αναλαμβάνει την παραγωγή προγεστερόνης και το κίτρινο σώμα αποικοδομείται σε λευκό σωμάτιο χωρίς απώλεια εμβρύου/εμβρύου.
Η ωχρινική υποστήριξη αναφέρεται στη χορήγηση φαρμάκων (γενικά προγεστινών) με σκοπό την αύξηση της επιτυχίας της εμφύτευσης και της πρώιμης εμβρυογένεσης, συμπληρώνοντας έτσι τη λειτουργία του ωχρού σωματίου.
Περιεκτικότητα σε καροτενοειδή
ΕπεξεργασίαΤο κίτρινο χρώμα και το όνομα του ωχρού σωματίου, όπως αυτό της ωχράς κηλίδας του αμφιβληστροειδούς, οφείλεται στη συγκέντρωσή του σε ορισμένα καροτενοειδή, ιδιαίτερα στη λουτεΐνη. Το 1968, μια έκθεση έδειξε ότι η β-καροτίνη συντέθηκε σε εργαστηριακές συνθήκες σε φέτες ωχρού σωματίου από αγελάδες. Ωστόσο, έχουν γίνει προσπάθειες να επαναληφθούν αυτά τα ευρήματα, αλλά δεν έχουν πετύχει.[12] Η ιδέα δεν είναι προς το παρόν αποδεκτή από την επιστημονική κοινότητα. Μάλλον, το ωχρό σωμάτιο συγκεντρώνει καροτενοειδή από τη διατροφή του θηλαστικού.
Πρόσθετες εικόνες
Επεξεργασία-
 Σειρά αλλαγών στην ωοθήκη
Σειρά αλλαγών στην ωοθήκη -
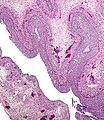 Ωχρινοθυλακιώδης κύστη. Χρώση H&E.
Ωχρινοθυλακιώδης κύστη. Χρώση H&E.


Παραπομπές
Επεξεργασία- ↑ «Histology Laboratory Manual». www.columbia.edu. Αρχειοθετήθηκε από το πρωτότυπο στις 6 Μαΐου 2017. Ανακτήθηκε στις 3 Μαΐου 2018.
- ↑ Inquiry Into Biology. McGraw-Hill Ryerson. 2007. σελίδες 497. ISBN 978-0-07-096052-7.
- ↑ Karch, σελ. 657.
- ↑ «FSH and follucogenesis: from physiology to ovarian stimulation». Reproductive biomedicine Online. 2006. Αρχειοθετήθηκε από το πρωτότυπο στις 15 Ιουλίου 2011. Ανακτήθηκε στις 26 Μαΐου 2009.
- ↑ 5,0 5,1 5,2 5,3 5,4 Boron, σελ. 1300.
- ↑ Niswender GD (March 2002). «Molecular control of luteal secretion of progesterone». Reproduction 123 (3): 333–9. doi:. PMID 11882010.
- ↑ 7,0 7,1 7,2 «Antioxidant capacity is correlated with steroidogenic status of the corpus luteum during the bovine estrous cycle». Biochim. Biophys. Acta 1380 (1): 133–40. March 1998. doi:. PMID 9545562. https://zenodo.org/record/890705.
- ↑ «Electron leakage from the mitochondrial NADPH-adrenodoxin reductase-adrenodoxin-P450scc (cholesterol side chain cleavage) system». Arch. Biochem. Biophys. 305 (2): 489–98. September 1993. doi:. PMID 8396893. https://zenodo.org/record/890721.
- ↑ «Electron leakage from the adrenal cortex mitochondrial P450scc and P450c11 systems: NADPH and steroid dependence». Arch. Biochem. Biophys. 317 (2): 412–6. March 1995. doi:. PMID 7893157. https://zenodo.org/record/890751.
- ↑ 10,0 10,1 10,2 The IUPS Physiome Project --> Female Reproductive System – Cells Αρχειοθετήθηκε 2009-12-10 στο Wayback Machine. Retrieved on Nov 9, 2009
- ↑ Marieb, Elaine (2013). Anatomy & physiology. Benjamin-Cummings. σελ. 915. ISBN 9780321887603.
- ↑ Brian Davis. Carotenoid metabolism as a preparation for function. Pure and Applied Chemistry, Vol. 63, No. 1, pp. 131–140, 1991. available online. Αρχειοθετήθηκε 2011-07-26 στο Wayback Machine. Accessed April 30, 2010.
Βιβλιογραφία
Επεξεργασία- Karch, Amy (2017). Focus on nursing pharmacology. Philadelphia: Wolters Kluwer. ISBN 9781496318213.
- Boron, Walter (2005). Medical physiology : a cellular and molecular approach. Philadelphia, Penns: Elsevier Saunders. ISBN 1-4160-2328-3.